
notokord - Notochord
| notokord | |
|---|---|
 Kırk beş saatlik kuluçkalık bir civciv embriyosunun enine kesiti.
| |
| Detaylar | |
| haberci | eksenel mezoderm |
| yol açar | çekirdek pulposus |
| tanımlayıcılar | |
| Latince | notokorda |
| ağ | D009672 |
| TE | E5.0.1.1.0.0.8 |
| Anatomik terminoloji | |
Olarak anatominin , sırt ipliği için benzer bir malzemeden oluşturulan bir esnek çubuk kıkırdak . Bir türün yaşam döngüsünün herhangi bir aşamasında bir notokord varsa , bu tanım gereği bir kordalıdır . Notokord ön-arka eksen boyunca uzanır ( önden arkaya ), genellikle dorsalde embriyonun ventral yüzeyinden daha yakındır ve mezodermden türetilen hücrelerden oluşur .
Notokordun en yaygın olarak belirtilen işlevleri şunlardır: gelişim sırasında çevreleyen dokuya yön sinyalleri sağlayan bir orta hat dokusu olarak, bir kas bağlanma yeri olarak ve bir omur öncüsü olarak.
Gelen lancelets Notokordun vücudun ana yapısal destek olarak hayatı boyunca devam eder. Olarak tunisatlar Notokordun yetişkin bir hayvanda tamamen yok olma, ancak larva aşamasında mevcut bulunmaktadır. Toplamda omurgalıların dışında hagfish , sırt ipliği içine geliştirir omurgadan haline vertebra ve intervertebral disklerin olan merkezi orijinal Notokordun benzer bir yapı kazanır.
Yapı
Notokord, bağırsağın dorsalinde ve nöral tüpün ventralinde gelişen uzun, çubuk benzeri bir yapıdır. Notokord, esas olarak, iki karşıt sarmal halinde sarılmış bir kolajen lifleri kılıfıyla kaplanmış bir glikoprotein çekirdeğinden oluşur . Glikoproteinler, vakuollü, şişkin hücrelerde depolanır. Bu lifler arasındaki açı, çekirdekteki artan basıncın, uzama ve incelmeye karşı kısalma ve kalınlaşma ile sonuçlanıp sonuçlanmayacağını belirler.
Bu kas liflerinin kasılması, yüzmeye benzer bir yan yana hareketle sonuçlanır. Sertleştirilmiş notokord, solucanınki gibi teleskopik hareket yoluyla hareketi engeller .
Sinyalizasyon ve geliştirmede rol
Notokord, gelişimin işaretlenmesinde ve koordine edilmesinde önemli bir rol oynar. Modern omurgalıların embriyoları gastrulasyon sırasında geçici notokord yapıları oluşturur . Notokordun bulunan ventral için nöral tüp .
Notogenesis tarafından Notokordun gelişmesidir epiblasts zeminini oluşturan amniyon boşluğu. Progenitör notokord, ilkel düğüm ve çukurdan göç eden hücrelerden türetilir . Notokord gastrulasyon sırasında oluşur ve kısa bir süre sonra nöral tüpün gelişimini senkronize ederek nöral plakanın ( nörülasyon ) oluşumunu indükler . Açık ventral sinir oluğun yönünde bir eksenel koyulaştırıcı endoderm gerçekleşir. (İki ayaklı kordalılarda, örneğin insanlarda, bu yüzeye uygun şekilde ön yüzey denir ). Bu kalınlaşma, kenarları anastomoz olan (temas eden) bir oluk (kordal oluk) olarak görünür ve böylece onu daha sonra endodermden ayrılan çokgen şekilli hücrelerden (notokord) oluşan katı bir çubuğa dönüştürür.
Omurgalılar, bu gelecek vertebral kolonun tüm uzunluğu boyunca uzanır ve uzak ön ucuna kadar ulaşır orta beynin gelecekteki bölgesinde bir kanca benzeri ekstremitenin biter, sırtı sella'nın ait sfenoid kemiğin . Başlangıçta, nöral tüp ile yolk kesesinin endodermi arasında bulunur; yakında, notokord , medial olarak büyüyen ve onu çevreleyen mezoderm tarafından onlardan ayrılır . Nöral tüp ve notochord çevreleyen mezodermden, kafatası , omurga ve membranlar arasında beyin ve medulla spinalis geliştirilmektedir. İlkel düğümden kaynaklandığı ve nihayetinde mezodermal boşlukla konumlandığı için mezodermden türetildiği kabul edilir.
Notokordun postembriyonik bir kalıntısı , intervertebral disklerin nükleus pulposusunda bulunur. İzole edilmiş notokordal kalıntılar, nükleus pulposus'taki soy spesifik hedeflerinden kaçabilir ve bunun yerine , notokord hücrelerinin büyük ölçüde gerilediği vertebra gövdelerinin dış yüzeylerine yapışabilir.
Amfibiler ve balıklarda
Amfibilerin ve balıkların gelişimi sırasında, notokord , vasküler endotelyal büyüme faktörünün salgılanması yoluyla hipokordun gelişimini indükler . Hipokord, notokord'un ventralindeki geçici bir yapıdır ve dorsal aortun doğru gelişiminden birincil derecede sorumludur.
İnsanlarda
4 yaşına gelindiğinde, tüm notokord kalıntısı, kökeni belirsiz bir kondrosit benzeri hücre popülasyonu ile değiştirilir . Omurdaki notokord hücrelerinin kalıcılığı patolojik bir duruma neden olabilir: kalıcı notokord kanalı . Embriyonik gelişim sırasında notokord ve nazofarenks düzgün bir şekilde ayrılmazsa, bir depresyon (Tornwaldt bursa) veya Tornwaldt kisti oluşabilir. Hücreler, kordoma adı verilen nadir bir kanserin muhtemel öncüleridir .
Nöroloji
Notokord üzerine yapılan araştırmalar, merkezi sinir sisteminin gelişimini anlamada önemli bir rol oynamıştır . Normal notokord yerinin 180 derece zıddı olan dorsal nöral tüpün yakınında ikinci bir notokord naklederek ve ifade ederek, dorsal tüpte motor nöronların oluşumunu indükleyebilir . Motor nöron oluşumu genellikle ventral nöral tüpte meydana gelirken, dorsal tüp genellikle duyu hücrelerini oluşturur .
Notokord, organogenezi düzenleyen ve motor nöronların gelişiminin sinyalleşmesinde kritik bir role sahip olan anahtar bir morfojen olan sonik kirpi (SHH) adı verilen bir protein salgılar . Notokord tarafından SHH salgılanması, gelişmekte olan embriyoda dorsal-ventral eksenin ventral kutbunu oluşturur.
Kordalılarda Evrim

Notokord , kordalıların tanımlayıcı özelliğidir ( sinapomorfi ) ve en eski kordalıların çoğunda yaşam boyunca mevcuttu. Her ne kadar stomochord ait yarı sırtipliler kez homolog olduğu düşünülüyordu, artık bir olarak görülüyor yakınlaşma . Pikaia'nın proto-Notokordun sahip gibi görünmektedir, ve notokorda gibi çeşitli bazal omurgalılar içinde mevcut olan Haikouella , Haikouichthy'dir ve Myllokunmingia , tüm Kambriyen'de .
Ordovisien okyanuslar birçok farklı türler dahil Agnatha ve erken gerçekçeneliler ya bağlı kemik elemanları ile veya özellikle olmadan, notokorda sahip Konodontlar , zırhlı balıklar ve ostracoderms . Hatta vertebral kolonun evrimi sonra kıkırdaklı ve kemikli balıkların bu takson ortak kalmıştır ve iyi fosiller kayıtlarında temsil edilir. Bazı türler (aşağıdaki listeye bakınız) ilkel duruma geri dönerek notokordları yetişkinliğe kadar korumuştur, ancak bunun nedenleri iyi anlaşılmamıştır.
Notokord'un evrimsel kökenine ilişkin senaryolar, Annona, Holland ve D'Aniello (2015) tarafından kapsamlı bir şekilde gözden geçirilmiştir. Bu fikirlerin birçoğunun moleküler filogenetik ve gelişimsel genetikteki ilerlemeler tarafından iyi bir şekilde desteklenmemesine rağmen, ikisinin aslında modern moleküler yaklaşımların teşviki altında yeniden canlandırıldığına dikkat çekiyorlar (birincisi, notokord'un kordalılarda de novo evrimleştiğini öne sürüyorlar. ve ikincisi onu, kordalıların annelid benzeri atalarında bulunan homolog bir yapı olan aksokordan türetir). Bu iki senaryo (veya muhtemelen henüz önerilecek başka bir senaryo) arasında karar vermek, geniş bir hayvan yelpazesinde gen düzenleyici ağların çok daha kapsamlı çalışmalarıyla kolaylaştırılmalıdır.
Post-embriyonik tutma
Çoğu omurgalıda, notokord ikincil yapılara dönüşür. Diğer kordatlarda , notokord, temel bir anatomik yapı olarak korunur. Chordata filumu içindeki notokord evrimi Holland ve Somorjai (2020) tarafından ayrıntılı olarak ele alınmıştır.
Aşağıdaki organizmalar post-embriyonik notokord tutar:
- Acipenseriformes ( kürek balığı ve mersin balığı )
- Lancelet ( Amphioxus'un )
- Tunikat (yalnızca larva evresi)
- Hagfish
- Lamprey
- Coelacanth
- Afrika akciğer balığı
- iribaşlar
- Ostrakodermler (soyu tükenmiş)
Amphioxus içinde
Lancelet'in notokord, nöral tüpün ön ucunun ötesine uzanır. Bu çıkıntı, hayvanın sığ suların tortusu içinde yuva yapmasına izin vererek ikinci bir amaca hizmet eder. Orada, amphioxus bir filtre besleyicidir ve ömrünün çoğunu kısmen tortu içinde geçirir.
Ek resimler
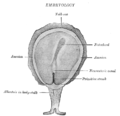
Concolor gibbon ( Hylobates concolor ) embriyosunun yüzey görünümü .
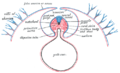
Civcivde amniyonun oluşum şeklini gösteren enine kesit diyagramı.

Arka beyin bölgesinde, yaklaşık on iki günlük bir insan embriyosunun kafasından kesit.
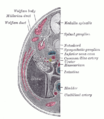
Sekiz buçuk ila dokuz haftalık insan embriyosunun enine kesiti.




Referanslar
- ^ a b Stemple, Derek L. (2005-06-01). "Notokordun yapısı ve işlevi: kordat için gerekli bir organ" . Geliştirme . 132 (11): 2503–2512. doi : 10.1242/dev.01812 . ISSN 0950-1991 . PMID 15890825 .
- ^ Krämer, Jürgen (2009). Intervertebral Disk Hastalıkları: Nedenleri, Tanı, Tedavi ve Profilaksi . Thieme. s. 15-17. ISBN'si 978-3-13-582403-1.
- ^ MAR Köhl (2000). "Elyaf Sarımlı Hidrolik İskeletlerin Mekanik Tasarımı: Embriyonik Notokordların Sertleşmesi ve Doğrultulması" . Amerikalı Zoolog . 40 : 28–041. doi : 10.1093/icb/40.1.28 .
- ^ a b Homberger, Dominique G. (2004). Omurgalı diseksiyonu . Walker, Warren F. (Warren Franklin), Walker, Warren F. (Warren Franklin). (9. baskı). Belmont, CA: Thomson Brooks/Cole. ISBN'si 0-03-022522-1. OCLC 53074665 .
- ^ "Üç katmanlı mikrop diski (3. hafta)" . www.embriyoloji.ch .
- ^ Hood, Rousseaux, Blakley, Ronald D., Colin G., Patricia M. (29 Mayıs 2007). "Embriyo ve Fetüs". Toksikolojik Patoloji El Kitabı (İkinci Baskı) . Academic Press, Elsevier Inc. tarafından yayınlandı. 2 : 895–936. doi : 10.1016/b978-0-12-330215-1.50047-8 . ISBN'si 9780123302151.CS1 bakımı: birden çok ad: yazar listesi ( bağlantı )
- ^ Henry Gray (1918). İnsan Vücudunun Anatomisi . Lea ve Febiger. s. 52–54.
- ^ Gary C. Schoenwolf; Steven B. Bleyl; Philip R. Brauer; Philippa H. Francis-West (1 Aralık 2014). Larsen'in İnsan Embriyolojisi E-Kitabı . Elsevier Sağlık Bilimleri. s. 71–72. ISBN'si 978-1-4557-2791-9.
- ^ Choi, K.; Cohn, Martin J.; Harfe, Brian D. (2009). "Farede Nucleus Pulposus Prekürsör Hücreleri ve Notokord Kalıntılarının Belirlenmesi: Disk Dejenerasyonu ve Kordoma Oluşumunun Etkileri" . Gelişim Dinamikleri . 237 (12): 3953-3958. doi : 10.1002/dvdy.21805 . PMC 2646501 . PMID 19035356 .
- ^ Cleaver, Ondine (2000). "Notokord tarafından endoderm desenleme: Xenopus'ta hipokordun gelişimi" (PDF) . Geliştirme . 127 (4): 869-979. doi : 10.1242/dev.127.4.869 . PMID 10648245 .
- ^ Urban, JPG (2000). "Gelişimden Dejenerasyona İntervertebral Diskin Çekirdeği" . Bütünleştirici ve Karşılaştırmalı Biyoloji . 40 : 53–061. doi : 10.1093/icb/40.1.53 .
- ^ Christopherson, Lr; Rabin, Bm; Hallam, Dk; Russell, Ej (1 Ocak 1999). "Notokord kanalının kalıcılığı: MR ve düz film görünümü" (Serbest tam metin) . Amerikan Nöroradyoloji Dergisi . 20 (1): 33–6. ISSN 0195-6108 . PMID 9974055 .
- ^ Moody MW, Chi DH, Chi DM, Mason JC, Phillips CD, Gross CW, et al. (2007). "Tornwaldt kisti: insidans ve bir olgu sunumu" . Kulak Burun Boğaz J . 86 (1): 45–7, 52. doi : 10.1177/014556130708600117 . PMID 17315835 .
- ^ Pillai S, Govender S (2018). "Sakral kordoma: Literatürün gözden geçirilmesi" . J Ortop . 15 (2): 679-684. doi : 10.1016/j.jor.2018.04.001 . PMC 5990241 . PMID 29881220 .
- ^ Echelard, Y; Epstein, Dj; St-Jacques, B; Shen, L; Mohler, J; Mcmahon, Ja; Mcmahon, Ap (Aralık 1993). "Varsayılan sinyal molekülleri ailesinin bir üyesi olan sonik kirpi, CNS polaritesinin düzenlenmesinde rol oynar" . Hücre . 75 (7): 1417–30. doi : 10.1016/0092-8674(93)90627-3 . PMID 7916661 . S2CID 6732599 .
- ^ Kardong, Kenneth V. (1995). Omurgalılar: karşılaştırmalı anatomi, işlev, evrim . McGraw-Hill. s. 55, 57. ISBN 978-0-697-21991-6.
- ^ "Arşivlenmiş kopya" . Arşivlenmiş orijinal 2006-03-13 tarihinde . 2007-09-05 alındı .CS1 bakımı: başlık olarak arşivlenmiş kopya ( bağlantı )
- ^ "Arşivlenmiş kopya" . Arşivlenmiş orijinal 20 Aralık 2010 tarihinde . Erişim tarihi: 21 Kasım 2009 .CS1 bakımı: başlık olarak arşivlenmiş kopya ( bağlantı )
- ^ Annana, G.; Hollanda, ND; D'Aniello, S. (2015). "Notokord Evrimi" . EvoDevo . 6 . makale 30. doi : 10.1186/s13227-015-0025-3 . PMC 4595329 . PMID 26446368 .
- ^ Hollanda, Kuzey Kore; Somorjai, IML (2020). "Seri blok yüz SEM, kök hücrelerin bir omurgasız kordalı Bahamalar neşterinde yetişkin notokord büyümesine katılabileceğini öne sürüyor" . EvoDevo . 11 . madde 22. doi : 10.1186/s13227-020-00167-6 .
- ^ Joseph J. Luczkovich; Philip J.Motta; Stephen F. Norton; Karel F. Liem (17 Nisan 2013). Balıkların ekomorfolojisi . Springer Bilim ve İş Medyası. P. 201. ISBN'si 978-94-017-1356-6.